Results
Here are the results for simulated and real data.
Note
retroCNVs - polymorphic retrocopies
Simulated data
Our dataset for testing is composed of 100 simulated human whole-genome sequencing with 20x of depth and in average 30 randomly distributed retrocopies each. Simulation with low coverage of (‘only’) 20x in sequencing depth (i.e., heterozygotic events have only 10x coverage). This strategy allowed us to check the capability of sideRETRO to identify retroCNVs events even in a “non-ideal scenario” of low sequencing coverage. In total, we had a list of 100 retrocopies consisting of the last 1000 bases of the largest transcript of the parental gene - which were randomly raflled as well. All retrocopies was stochastically designed for chromosome, position, strand and zygosity.
The simulated retrocopy data is composed of three sets of retroCNVs events:
fixed or highly frequent events;
polymorphic events (shared by some of the simulated genomes);
iii) somatic events (in only one genome) in simulation. It allowed us to check sideRETRO performance for these different types of retroCNVs.
Simulation
We developed a pipeline, which randomly generates our simulated dataset and
make some analysis of performance. All scripts can be downloaded at
simulation.tar.gz. We used the
SANDY tool (version v0.23), A straightforward and complete next-generation
sequencing read simulator [2], for simulate all 100 genomes according to the
structural variations that we designed and according to the sampling. We used
the reference human genome v38 and the GENCODE annotation v32.
REF_FASTA=/assets/hg38.fa
PC_FASTA=/assets/gencode.v32.pc_transcripts.fa
COHORT=100
RTC_NUM=100
LEN=1000
DEPTH=20
SANDY_SEED=1
SEED=17
# Genearte sequences
scripts/catch \
--seed=$SEED \
--rtc_num=$RTC_NUM \
--length=$LEN \
"$PC_FASTA" > rtc_100.tsv
# Build our cohort
scripts/build \
--cohort=$COHORT \
--seed=$SEED \
--output-dir=build \
"$REF_FASTA" \
rtc_100.tsv
# Retrocopies by individual
IND=($(ls build/*.sandy))
# Load build values to SANDY
for ind in "${IND[@]}"; do
sandy variation add \
--structural-variation=$(basename $ind '.sandy') \
$ind
done
mkdir -p sim
# Simulate all genomes
for ind in "${IND[@]}"; do
sandy_index=$(basename $ind '.sandy')
sandy genome \
--id='%i.%U_%c:%S-%E_%v' \
--structural-variation=$sandy_index \
--output-dir="sim/$sandy_index" \
--jobs=20 \
--seed=$SANDY_SEED \
--quality-profile='hiseq_101' \
--coverage=$DEPTH \
--verbose \
$REF_FASTA
done
As result we have a pair of FASTQ files (forward and reverse complement) for each simulated individual. Next it is required to align our sequencing data against the human reference genome in order to generate mapped files in SAM format. We used BWA aligner (version 0.7.9) [3] for accomplish this task.
# Individual directories with the
# simulated data
IND_DIR=($(ls -d sim/*))
# Reference genome
REF_FASTA="/assets/hg38.fa"
# Index reference genome
bwa index $REF_FASTA
mkdir -p align
# Alignment
for ind in "${IND[@]}"; do
id="$(basename $ind)"
bwa mem \
-t 10 \
$REF_FASTA \
$ind/out_R1_001.fastq.gz \
$ind/out_R2_001.fastq.gz > "align/$id.sam"
done
After our simulated dataset was ready, we run sideRETRO v0.14.1:
# Our simulated SAM files list
LIST=($(ls align/*.sam))
# GENCODE annotation v32
ANNOTATION=/assets/gencode.v32.annotation.gff3
# GENCODE reference genome
REF_FASTA=/assets/hg38.fa
# Run process-sample step
sider process-sample \
--prefix=sim \
--cache-size=20000000 \
--output-dir=sider \
--threads=20 \
--alignment-frac=0.9 \
--phred-quality=20 \
--sorted \
--log-file=ps.log \
--annotation-file=$ANNOTATION \
"${LIST[@]}"
# Run merge-call step
sider merge-call \
--cache-size=20000000 \
--epsilon=500 \
--min-pts=10 \
--log-file=mc.log \
--threads=20 \
--phred-quality=20 \
--in-place \
sider/sim.db
# Finally run make-vcf
sider make-vcf \
--log-file=vcf.log \
--reference-file=$REF_FASTA \
--prefix=sim \
--output-dir=sider \
sider/sim.db
Finally, with the sideRETRO’s VCF made, we analysed the performance:
# Generate comparations for analysis
scripts/compare sider/sim.vcf build
# Confusion analysis
scripts/confusion analysis > confusion.tsv
# Just a look
$ column -t confusion.tsv | head
IND TP FP FN PPV/Precision TPR/Recall F1-score
analysis/ind0.tsv 38 0 9 1.000000 0.808511 0.894118
analysis/ind1.tsv 36 2 11 0.947368 0.765957 0.847059
analysis/ind2.tsv 33 1 10 0.970588 0.767442 0.857143
analysis/ind3.tsv 35 1 12 0.972222 0.744681 0.843373
analysis/ind4.tsv 29 1 9 0.966667 0.763158 0.852941
analysis/ind5.tsv 37 4 12 0.902439 0.755102 0.822222
analysis/ind6.tsv 45 0 10 1.000000 0.818182 0.900000
analysis/ind7.tsv 37 2 11 0.948718 0.770833 0.850575
analysis/ind8.tsv 32 2 11 0.941176 0.744186 0.831169
Analysis
Parental Gene |
SIMULATED |
FOUND (79 events) |
|||||
|---|---|---|---|---|---|---|---|
Chr |
Position |
Pol |
LINE/SINE |
Chr |
Position |
Pol |
|
ALG2 |
chr10 |
30778982 |
- |
N |
chr10 |
30778981 |
- |
ARMC2 |
chr5 |
52723637 |
- |
Y |
chr5 |
52723638 |
- |
ATG2B |
chr5 |
177026995 |
- |
N |
chr5 |
177026990 |
- |
BTF3 |
chr7 |
146774631 |
- |
N |
chr7 |
146774629 |
- |
C2orf92 |
chr6 |
112158328 |
- |
N |
chr6 |
112158327 |
- |
C8orf76 |
chr9 |
94927085 |
- |
N |
chr9 |
94927084 |
- |
C9orf64 |
chr17 |
40139106 |
+ |
Y |
chr17 |
40139104 |
+ |
CABP7 |
chr5 |
153788597 |
+ |
Y |
chr5 |
153788596 |
+ |
CARD8 |
chrX |
99922659 |
+ |
N |
chrX |
99922658 |
+ |
CASTOR3 |
chr3 |
189081695 |
- |
N |
chr3 |
189081692 |
- |
CDH22 |
chr9 |
113306486 |
- |
Y |
chr9 |
113306485 |
- |
CFAP69 |
chr11 |
10733916 |
- |
N |
chr11 |
10733915 |
- |
COL4A3 |
chr16 |
46427444 |
+ |
N |
chr16 |
46427444 |
+ |
COPS2 |
chr1 |
38773310 |
- |
Y |
chr1 |
38773309 |
- |
CPNE7 |
chr9 |
42228417 |
+ |
Y |
chr9 |
42228469 |
. |
DENND2D |
chr18 |
37314709 |
+ |
N |
chr18 |
37314708 |
+ |
DNAJC27 |
chr12 |
60940050 |
- |
N |
chr12 |
60940049 |
- |
EPC2 |
chr13 |
94468157 |
- |
N |
chr13 |
94468156 |
- |
EPS8 |
chr21 |
26428011 |
+ |
N |
chr21 |
26428011 |
+ |
ERCC4 |
chr6 |
93262920 |
+ |
N |
chr6 |
93262919 |
+ |
FAAP20 |
chr9 |
77384901 |
- |
N |
chr9 |
77384898 |
- |
FAM177B |
chr12 |
130498191 |
+ |
N |
chr12 |
130498188 |
+ |
FAM71E2 |
chr2 |
225319689 |
+ |
N |
chr2 |
225319688 |
+ |
HAO2 |
chr14 |
69901152 |
+ |
N |
chr14 |
69901150 |
+ |
HEG1 |
chr3 |
15517386 |
- |
Y |
chr3 |
15517382 |
- |
HIP1 |
chr8 |
75177754 |
+ |
Y |
chr8 |
75177754 |
+ |
IL1R1 |
chr8 |
30386429 |
- |
N |
chr8 |
30386427 |
- |
IQGAP3 |
chr6 |
124358143 |
+ |
Y |
chr6 |
124358101 |
+ |
KIF7 |
chrX |
89251626 |
- |
Y |
chrX |
89251603 |
- |
LAMP1 |
chr13 |
87908197 |
- |
N |
chr13 |
87908197 |
- |
LARS |
chr9 |
64069435 |
+ |
Y |
chr9 |
64069377 |
+ |
LRRC6 |
chr4 |
180728002 |
- |
N |
chr4 |
180728002 |
- |
MACROD2 |
chr20 |
18178487 |
+ |
N |
chr20 |
18178486 |
+ |
MYH10 |
chr4 |
186290075 |
+ |
Y |
chr4 |
186290074 |
+ |
MYH7B |
chr13 |
104241206 |
+ |
N |
chr13 |
104241205 |
+ |
MYO7A |
chr11 |
14072547 |
+ |
N |
chr11 |
14072546 |
+ |
NAE1 |
chr18 |
74528384 |
+ |
Y |
chr18 |
74528383 |
+ |
OR14A16 |
chr1 |
52758590 |
+ |
N |
chr1 |
52758589 |
+ |
OR51M1 |
chr2 |
37409208 |
- |
N |
chr2 |
37409207 |
- |
OSER1 |
chr5 |
53846631 |
- |
Y |
chr5 |
53846596 |
- |
PAFAH1B1 |
chr15 |
86208543 |
+ |
Y |
chr15 |
86208562 |
+ |
PDGFB |
chr8 |
133462380 |
- |
N |
chr8 |
133462379 |
- |
PFKFB2 |
chr5 |
36822019 |
- |
N |
chr5 |
36822019 |
- |
PLCB1 |
chr9 |
25165703 |
+ |
Y |
chr9 |
25165702 |
+ |
PNRC1 |
chr15 |
48607415 |
+ |
N |
chr15 |
48607414 |
+ |
PRMT2 |
chr8 |
50511539 |
- |
Y |
chr8 |
50511540 |
- |
PRPF18 |
chr20 |
51460729 |
+ |
Y |
chr20 |
51460728 |
+ |
PRSS45P |
chr19 |
5420707 |
- |
Y |
chr19 |
5420706 |
- |
PTPRF |
chr19 |
7227546 |
+ |
Y |
chr19 |
7227546 |
+ |
RAB18 |
chr4 |
10281361 |
- |
N |
chr4 |
10281361 |
- |
RAB5B |
chr6 |
46561322 |
+ |
N |
chr6 |
46561322 |
+ |
RADX |
chr12 |
117277769 |
+ |
N |
chr12 |
117277768 |
+ |
RASGEF1C |
chr5 |
115992817 |
+ |
N |
chr5 |
115992816 |
+ |
RBM4 |
chr7 |
101199285 |
+ |
Y |
chr7 |
101199284 |
+ |
RMDN3 |
chr3 |
28655572 |
- |
N |
chr3 |
28655571 |
- |
RNF6 |
chr4 |
39797761 |
- |
Y |
chr4 |
39797759 |
- |
SART1 |
chr2 |
109317943 |
+ |
N |
chr2 |
109317942 |
+ |
SDHA |
chr4 |
179658356 |
+ |
N |
chr4 |
179658355 |
+ |
SEZ6L |
chr18 |
560651 |
- |
Y |
chr18 |
560650 |
- |
SKP2 |
chr5 |
88746051 |
- |
N |
chr5 |
88746050 |
- |
SLC9A3 |
chr4 |
140369141 |
- |
N |
chr4 |
140369139 |
- |
SMTNL2 |
chr3 |
144112843 |
- |
N |
chr3 |
144112842 |
- |
SNRNP27 |
chrX |
13251389 |
- |
N |
chrX |
13251387 |
- |
STK17B |
chrX |
36995058 |
- |
Y |
chrX |
36995057 |
- |
TACO1 |
chrY |
12987416 |
+ |
Y |
chrY |
12987415 |
+ |
TMEM63C |
chr17 |
49131966 |
+ |
Y |
chr17 |
49131965 |
+ |
TMEM95 |
chr2 |
234301985 |
- |
Y |
chr2 |
234301984 |
- |
TSFM |
chr12 |
80384739 |
- |
Y |
chr12 |
80384736 |
- |
TUBGCP2 |
chr1 |
197233691 |
+ |
N |
chr1 |
197233690 |
+ |
VIPAS39 |
chr12 |
54021508 |
- |
N |
chr12 |
54021507 |
- |
WDR74 |
chr11 |
112552782 |
- |
N |
chr11 |
112552781 |
- |
WDR75 |
chr6 |
132636317 |
+ |
Y |
chr6 |
132636316 |
+ |
ZNF136 |
chr16 |
59509103 |
+ |
Y |
chr16 |
59509104 |
+ |
ZNF326 |
chr8 |
29273486 |
- |
Y |
chr8 |
29273482 |
- |
ZNF385A |
chr12 |
92752469 |
- |
N |
chr12 |
92752468 |
- |
ZNF431 |
chr16 |
88101015 |
- |
N |
chr16 |
88101015 |
- |
ZNF585A |
chr18 |
78888223 |
- |
Y |
chr18 |
78888222 |
- |
ZNF738 |
chr6 |
139608184 |
- |
N |
chr6 |
139608183 |
- |
ZNF793 |
chr9 |
120420222 |
+ |
N |
chr9 |
120420223 |
+ |
RetroCNV events not found by sideRetro (21 events) |
|||||||
Duplicated region |
|||||||
AC002310.4 |
chr9 |
94545202 |
- |
N |
chr8:115819078-115819180 |
||
AC135178.3 |
chr7 |
74794901 |
- |
N |
chr7:75151009-75151108 |
||
ACSBG2 |
chr21 |
43058887 |
- |
N |
chr21:6450515-6450614 |
||
ADD2 |
chr3 |
9759497 |
+ |
N |
No |
||
AL645922.1 |
chr6 |
38626680 |
- |
N |
No |
||
C21orf91 |
chr14 |
54886570 |
- |
Y |
Duplications: 7x genome |
||
CERS1 |
chr20 |
41341204 |
+ |
N |
No |
||
CWC25 |
chr13 |
39475646 |
- |
N |
No |
||
DHRSX |
chr5 |
166496220 |
- |
Y |
Highly repetitive region |
||
LETM1 |
chrY |
24793930 |
- |
N |
8 identical region in chrY |
||
MALL |
chr7 |
110598366 |
+ |
N |
No |
||
MRPS7 |
chr2 |
1490696 |
+ |
N |
chr2_KI270774v1_alt |
||
MTNR1A |
chr8 |
86938090 |
- |
N |
chrX, chr4 |
||
NDUFA6 |
chr10 |
38060463 |
+ |
N |
chr10:42588649-42588750 |
||
PLAC8 |
chr9 |
39225441 |
+ |
Y |
chr9:61393599-61393698 |
||
PTCHD4 |
chr15 |
31035142 |
- |
Y |
chr15_KI270905v1_alt |
||
SLC44A4 |
chrY |
4417954 |
+ |
Y |
chrX:90835484-90835583 |
||
STON2 |
chrX |
468106 |
+ |
N |
chrY:468056-468155 |
||
TAF7 |
chr22 |
22384919 |
- |
N |
chr22_KI270875v1_alt |
||
TBC1D3F |
chr16 |
65760883 |
+ |
Y |
No |
||
TRIM40 |
chr5 |
45713519 |
+ |
N |
No |
||
RetroCNV type |
# of simulated events |
Found events |
% |
|---|---|---|---|
Common |
25 |
19 |
76 |
Polymorphic |
50 |
42 |
84 |
Somatic |
25 |
18 |
72 |
Ind |
TP |
FP |
FN* |
PPV |
TPR (|*) |
F1 (|*) |
|---|---|---|---|---|---|---|
0 |
38 |
0 |
9|5 |
1.00 |
0.81|0.88 |
0.89|0.94 |
1 |
36 |
2 |
11|7 |
0.95 |
0.77|0.84 |
0.85|0.89 |
2 |
33 |
1 |
10|6 |
0.97 |
0.77|0.85 |
0.86|0.90 |
3 |
35 |
1 |
12|5 |
0.97 |
0.74|0.88 |
0.84|0.92 |
4 |
29 |
1 |
9|5 |
0.97 |
0.76|0.85 |
0.85|0.91 |
5 |
37 |
4 |
12|5 |
0.90 |
0.76|0.88 |
0.82|0.89 |
6 |
45 |
0 |
10|6 |
1.00 |
0.82|0.88 |
0.90|0.94 |
7 |
37 |
2 |
11|5 |
0.95 |
0.77|0.88 |
0.85|0.91 |
8 |
32 |
2 |
11|5 |
0.94 |
0.74|0.86 |
0.83|0.90 |
9 |
33 |
3 |
11|5 |
0.92 |
0.75|0.87 |
0.83|0.89 |
10 |
34 |
1 |
9|5 |
0.97 |
0.79|0.87 |
0.87|0.92 |
11 |
37 |
2 |
12|5 |
0.95 |
0.76|0.88 |
0.84|0.91 |
12 |
30 |
1 |
10|5 |
0.97 |
0.75|0.86 |
0.85|0.91 |
13 |
43 |
3 |
11|5 |
0.93 |
0.80|0.90 |
0.86|0.91 |
14 |
38 |
0 |
10|6 |
1.00 |
0.79|0.86 |
0.88|0.93 |
15 |
31 |
1 |
8|5 |
0.97 |
0.79|0.86 |
0.87|0.91 |
16 |
30 |
4 |
13|6 |
0.88 |
0.70|0.83 |
0.78|0.86 |
17 |
39 |
1 |
9|5 |
0.98 |
0.81|0.89 |
0.89|0.93 |
18 |
37 |
0 |
10|5 |
1.00 |
0.79|0.88 |
0.88|0.94 |
19 |
39 |
1 |
10|6 |
0.98 |
0.80|0.87 |
0.88|0.92 |
20 |
39 |
2 |
12|6 |
0.95 |
0.76|0.87 |
0.85|0.91 |
21 |
42 |
3 |
12|5 |
0.93 |
0.78|0.89 |
0.85|0.91 |
22 |
39 |
0 |
10|6 |
1.00 |
0.80|0.87 |
0.89|0.93 |
23 |
41 |
2 |
10|5 |
0.95 |
0.80|0.89 |
0.87|0.92 |
24 |
43 |
1 |
8|5 |
0.98 |
0.84|0.90 |
0.91|0.93 |
25 |
41 |
0 |
9|6 |
1.00 |
0.82|0.87 |
0.90|0.93 |
26 |
43 |
0 |
10|6 |
1.00 |
0.81|0.88 |
0.90|0.93 |
27 |
34 |
0 |
10|5 |
1.00 |
0.77|0.87 |
0.87|0.93 |
28 |
38 |
4 |
14|7 |
0.90 |
0.73|0.84 |
0.81|0.87 |
29 |
36 |
1 |
11|6 |
0.97 |
0.77|0.86 |
0.86|0.91 |
30 |
47 |
3 |
11|5 |
0.94 |
0.81|0.90 |
0.87|0.92 |
31 |
43 |
3 |
12|5 |
0.93 |
0.78|0.90 |
0.85|0.91 |
32 |
38 |
0 |
11|5 |
1.00 |
0.78|0.88 |
0.87|0.94 |
33 |
34 |
1 |
12|6 |
0.97 |
0.74|0.85 |
0.84|0.91 |
34 |
35 |
4 |
12|6 |
0.90 |
0.74|0.85 |
0.81|0.88 |
35 |
43 |
2 |
10|6 |
0.96 |
0.81|0.88 |
0.88|0.91 |
36 |
41 |
2 |
11|6 |
0.95 |
0.79|0.87 |
0.86|0.91 |
37 |
38 |
1 |
11|6 |
0.97 |
0.78|0.86 |
0.86|0.92 |
38 |
34 |
1 |
9|5 |
0.97 |
0.79|0.87 |
0.87|0.92 |
39 |
39 |
0 |
8|5 |
1.00 |
0.83|0.89 |
0.91|0.94 |
40 |
35 |
1 |
9|5 |
0.97 |
0.80|0.88 |
0.88|0.92 |
41 |
33 |
1 |
9|5 |
0.97 |
0.79|0.87 |
0.87|0.92 |
42 |
39 |
1 |
11|7 |
0.98 |
0.78|0.85 |
0.87|0.91 |
43 |
37 |
4 |
13|7 |
0.90 |
0.74|0.84 |
0.81|0.87 |
44 |
39 |
4 |
13|6 |
0.91 |
0.75|0.87 |
0.82|0.89 |
45 |
35 |
3 |
11|6 |
0.92 |
0.76|0.85 |
0.83|0.89 |
46 |
31 |
0 |
9|5 |
1.00 |
0.78|0.86 |
0.87|0.93 |
47 |
36 |
0 |
10|5 |
1.00 |
0.78|0.88 |
0.88|0.94 |
48 |
40 |
3 |
11|6 |
0.93 |
0.78|0.87 |
0.85|0.90 |
49 |
34 |
1 |
10|5 |
0.97 |
0.77|0.87 |
0.86|0.92 |
50 |
41 |
4 |
13|6 |
0.91 |
0.76|0.87 |
0.83|0.89 |
51 |
34 |
0 |
9|5 |
1.00 |
0.79|0.87 |
0.88|0.93 |
52 |
36 |
3 |
12|5 |
0.92 |
0.75|0.88 |
0.83|0.90 |
53 |
39 |
2 |
11|5 |
0.95 |
0.78|0.89 |
0.86|0.92 |
54 |
47 |
0 |
10|6 |
1.00 |
0.82|0.89 |
0.90|0.94 |
55 |
36 |
1 |
12|5 |
0.97 |
0.75|0.88 |
0.85|0.92 |
56 |
40 |
2 |
12|6 |
0.95 |
0.77|0.87 |
0.85|0.91 |
57 |
41 |
1 |
9|5 |
0.98 |
0.82|0.89 |
0.89|0.93 |
58 |
40 |
0 |
10|5 |
1.00 |
0.80|0.89 |
0.89|0.94 |
59 |
34 |
3 |
11|6 |
0.92 |
0.76|0.85 |
0.83|0.88 |
60 |
35 |
2 |
10|5 |
0.95 |
0.78|0.88 |
0.85|0.91 |
61 |
38 |
1 |
9|5 |
0.97 |
0.81|0.88 |
0.88|0.93 |
62 |
30 |
1 |
8|5 |
0.97 |
0.79|0.86 |
0.87|0.91 |
63 |
38 |
4 |
13|6 |
0.90 |
0.75|0.86 |
0.82|0.88 |
64 |
43 |
2 |
10|5 |
0.96 |
0.81|0.90 |
0.88|0.92 |
65 |
46 |
1 |
10|6 |
0.98 |
0.82|0.88 |
0.89|0.93 |
66 |
41 |
1 |
10|6 |
0.98 |
0.80|0.87 |
0.88|0.92 |
67 |
37 |
2 |
9|5 |
0.95 |
0.80|0.88 |
0.87|0.91 |
68 |
44 |
5 |
13|6 |
0.90 |
0.77|0.88 |
0.83|0.89 |
69 |
36 |
0 |
9|5 |
1.00 |
0.80|0.88 |
0.89|0.94 |
70 |
42 |
4 |
14|7 |
0.91 |
0.75|0.86 |
0.82|0.88 |
71 |
44 |
3 |
14|7 |
0.94 |
0.76|0.86 |
0.84|0.90 |
72 |
41 |
3 |
13|6 |
0.93 |
0.76|0.87 |
0.84|0.90 |
73 |
34 |
1 |
9|5 |
0.97 |
0.79|0.87 |
0.87|0.92 |
74 |
42 |
1 |
10|5 |
0.98 |
0.81|0.89 |
0.88|0.93 |
75 |
37 |
3 |
11|5 |
0.93 |
0.77|0.88 |
0.84|0.90 |
76 |
34 |
2 |
9|5 |
0.94 |
0.79|0.87 |
0.86|0.91 |
77 |
37 |
3 |
10|5 |
0.93 |
0.79|0.88 |
0.85|0.90 |
78 |
38 |
0 |
8|5 |
1.00 |
0.83|0.88 |
0.90|0.94 |
79 |
40 |
2 |
9|5 |
0.95 |
0.82|0.89 |
0.88|0.92 |
80 |
35 |
0 |
9|5 |
1.00 |
0.80|0.88 |
0.89|0.93 |
81 |
40 |
1 |
10|6 |
0.98 |
0.80|0.87 |
0.88|0.92 |
82 |
41 |
2 |
11|7 |
0.95 |
0.79|0.85 |
0.86|0.90 |
83 |
39 |
2 |
11|6 |
0.95 |
0.78|0.87 |
0.86|0.91 |
84 |
40 |
3 |
10|6 |
0.93 |
0.80|0.87 |
0.86|0.90 |
85 |
36 |
4 |
12|5 |
0.90 |
0.75|0.88 |
0.82|0.89 |
86 |
37 |
4 |
13|6 |
0.90 |
0.74|0.86 |
0.81|0.88 |
87 |
32 |
2 |
11|5 |
0.94 |
0.74|0.86 |
0.83|0.90 |
88 |
42 |
2 |
12|7 |
0.95 |
0.78|0.86 |
0.86|0.90 |
89 |
34 |
1 |
9|5 |
0.97 |
0.79|0.87 |
0.87|0.92 |
90 |
41 |
2 |
10|5 |
0.95 |
0.80|0.89 |
0.87|0.92 |
91 |
45 |
0 |
9|6 |
1.00 |
0.83|0.88 |
0.91|0.94 |
92 |
39 |
2 |
8|5 |
0.95 |
0.83|0.89 |
0.89|0.92 |
93 |
39 |
2 |
11|6 |
0.95 |
0.78|0.87 |
0.86|0.91 |
94 |
34 |
3 |
12|5 |
0.92 |
0.74|0.87 |
0.82|0.89 |
95 |
44 |
4 |
11|5 |
0.92 |
0.80|0.90 |
0.85|0.91 |
96 |
36 |
1 |
9|5 |
0.97 |
0.80|0.88 |
0.88|0.92 |
97 |
39 |
2 |
10|5 |
0.95 |
0.80|0.89 |
0.87|0.92 |
98 |
48 |
0 |
9|6 |
1.00 |
0.84|0.89 |
0.91|0.94 |
99 |
40 |
0 |
10|6 |
1.00 |
0.80|0.87 |
0.89|0.93 |
Total |
3806 |
172 |
1051|551 |
0.96 |
0.78|0.87 |
0.86|0.91 |
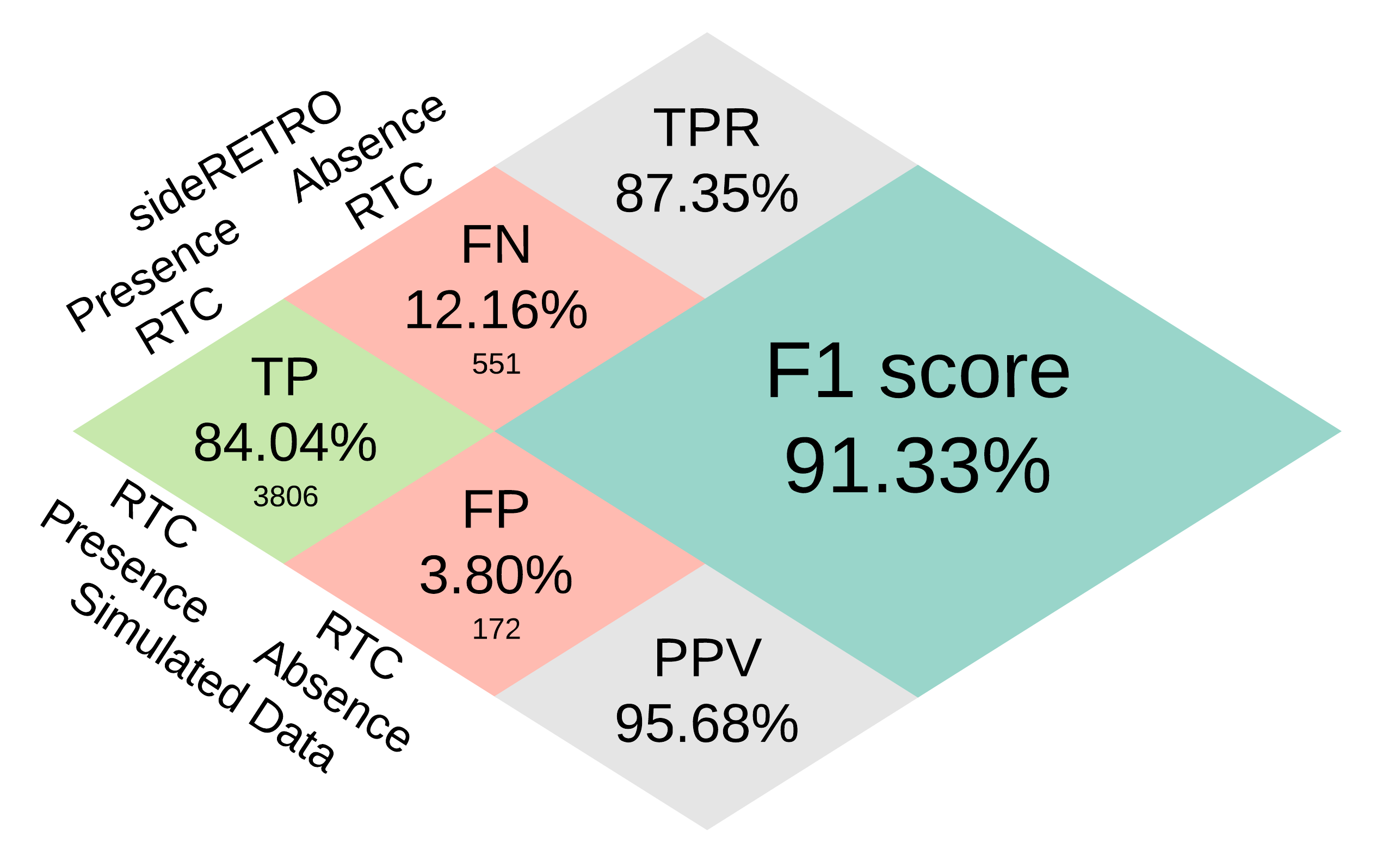
Overall performance for 86 simulated retroCNV events in mappeable genomic regions (Imbalanced confusion matrix). True Positive (TP), False Negative (FN), False Positive (FP), True Positive Rate or Recall (TPR), Positive Predictive Value or Precision (PPV) and F1 score.
Real data
The method developed and used by Abyzov et al. [1] relies on exon-exon junction reads to identify retroCNVs. In order to increase their candidate’s reliability, these authors performed experimental validations (Abyzov - Table 2). In summary, the authors. carried out PCR validation for nine putative retroCNVs and for six of them, they found their genomic insertion points (Red blocks). A retroCNV event is, by definition, a retroposition of an mRNA into a genomic region (i.e., it should have an insertion point, otherwise it could be a distinct retroCNV event, even from the same parental gene). Thus, in order to avoid misleading in data comparison, we selected those retroCNVs events validated by PCR and with a defined genomic insertion point.
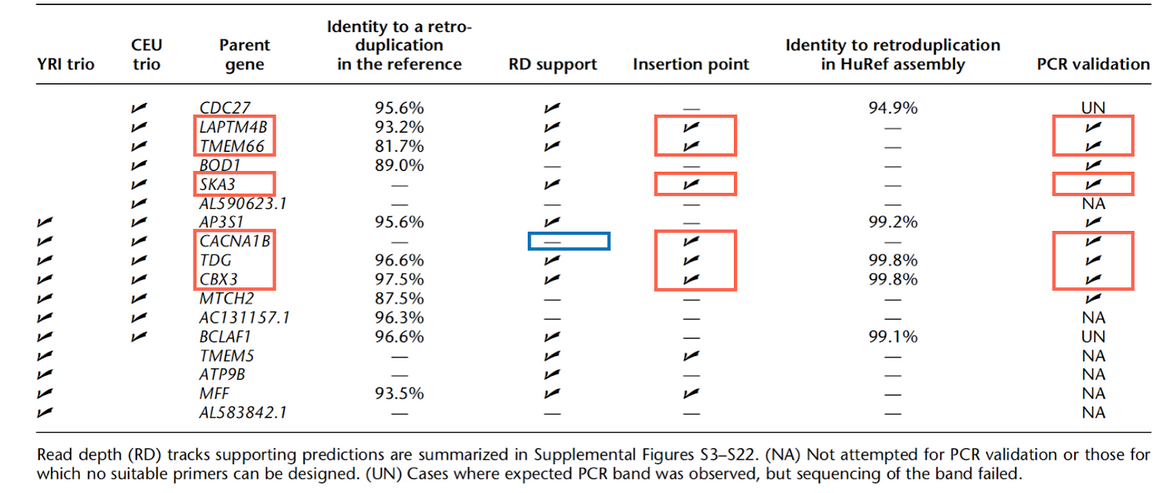
Highlighted in red: retroCNVs events presenting an insertion point and with PCR validation. Insertion point coordinates were retrieved from Table X, Abyzov et al, Genome Res, 2013.
Highlighted in blue: a lacking of read depth (RD) support to the candidate CACNA1B.
We called retroCNVs using the same 974 individuals from the fourteen (ASW, CEU, CHB, CHS, CLM, FIN, GBR, IBS, JPT, LWK, MXL, PUR, TSI, and YRI) 1000 Genome populations, which are reported in Supplementary _Table S1. Their six retroCNVs with PCR validation and a defined genomic insertion point (presented above, Abyzov - Table 2) were used. In summary, our pipeline (sideRETRO) identifies five (83.3%) and misses only one retroCNV (CACNA1B). Regarding the genotyping of retroCNVs shared by Abyzov and us, sideRETRO has a match of 70 genotyping out of 70 (100%), See tables below:
Parental Gene |
Insertion region (GRCh38; chromosome and position) |
|
|---|---|---|
Abyzov |
sideRETRO |
|
CBX3 |
15:40561954-40561998 |
15:40561980 |
LAPTM4B |
6:166920412-166920482 |
6:166920475 |
TMEM66* |
1:191829533-191829591 |
1:191829594 |
SKA3 |
11:108714998-108715054 |
11:108715020 |
TDG |
12:125316536-125316676 |
12:125316601 |
CACNA1B |
1:148027670-148027843 |
|
Parental Gene |
Populations |
|||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
ASW |
CEU |
CHB |
CHS |
CLM |
FIN |
GBR |
IBS |
JPT |
LWK |
MXL |
PUR |
TSI |
YRI |
|
CBX3 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
LAPTM4B |
0/0 |
1/1 |
0/0 |
0/0 |
1/1 |
1/1 |
1/1 |
0/0 |
0/0 |
0/0 |
0/0 |
1/1 |
1/1 |
0/0 |
TMEM66* |
0/0 |
1/1 |
0/0 |
0/0 |
0/0 |
1/1 |
1/1 |
0/0 |
0/0 |
0/0 |
0/0 |
1/1 |
1/1 |
0/0 |
SKA3 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
TDG |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
1/1 |
CACNA1B |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
1/0 |
Regarding the retroCNV event (parental gene CACNA1B; insertion region: chr1: 147499911-147500084) not identified by sideRETRO:
i) Curiously, Abyzov et al. did not find a good Read Depth Support for it (See above, marked in blue and in their manuscript);
ii) We found that its putative insertion region (GRCh37: chr1:147499911- 147500084; GRCh38: chr1:148,027,670-148,027,843) corresponds to a LTR region (Part A- below);
iii) This region has a second (quasi-perfect: only 2 mismatches) hit elsewhere, Part B;
iv) Moreover, this second hit is (suspiciously) near to a fixed retrocopy from the same parental gene, CACNA1B (Figure 1C). SideRETRO filters out retroCNVs (i.e., polymorphic) events inserted near a fixed retrocopy from the same parental gene, because they are usually results from false-positive alignments, since their likelihood of being real is very low (roughly = 1 / (genome size x number of genes; haploid genome: 3x109; the number of genes ~ 20k coding genes). Nevertheless, only a further experimental validation may confirm our hypothesis.
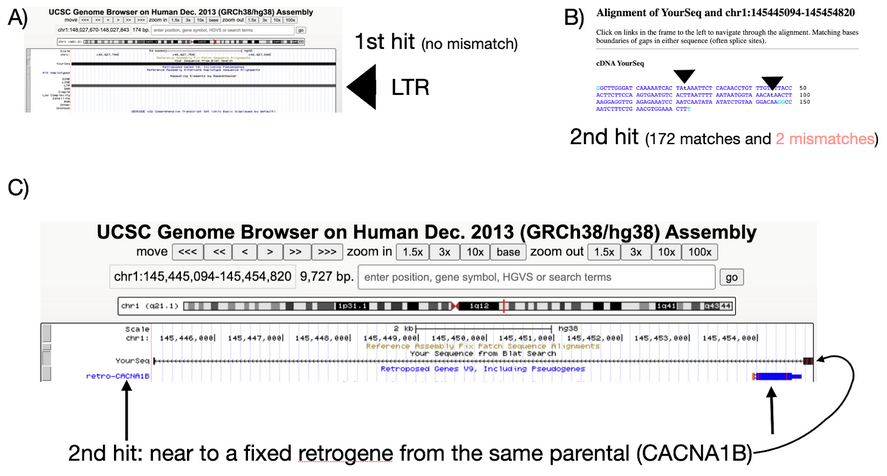
Genome alignment of the CACNA1B region defined by Abyzov et al. A) genomic alignment of the region defined as the insertion point of CACNA1B (in this case, GRCh38 was used). B) The second hit of this sequence into the genome (only two mismatcher in 174bp). C) The 2nd hit into the genome is near a fixed retrocopy from CACNA1B.
Thus, in summary, regarding the genotyping data, our pipeline presents a very good match ranging from 83.3% (considering all events) to 100% (excluding a “suspicious” candidate) against the experimental dataset from an independent group, Abyzov et al. (2013) Gen. Res.